从不确定性和熵到一致性和意识

从不确定性和熵到一致性和意识

CreateAMind
发布于 2026-03-11 17:57:03
发布于 2026-03-11 17:57:03
从不确定性和熵到一致性和意识
From uncertainty and entropy to coherence and consciousness


摘要 理解意识的神经基础仍是神经科学中的一个根本性挑战。本研究提出一个新框架,通过不确定性降低与负熵(negative entropy)的视角来概念化意识,并强调相干性在其涌现中的作用。感觉处理可能作为一种贝叶斯推理机制运作,旨在最小化大脑对外部刺激的不确定性;当不确定性降低至某一临界阈值以下时,意识觉知便会出现。在计算层面,这对应于信息不确定性的最小化;在物理层面,则对应于热力学熵的减少,从而将意识与负熵联系起来。本研究强调相干性在意识知觉中的作用,并通过探讨量子相干性与纠缠的潜在贡献,对现有模型(如整合信息理论)提出挑战。尽管目前尚缺乏直接的实证验证,我们提出一个假说:意识充当大脑的一种冷却机制,可通过神经回路的温度加以测量。这一观点为意识体验的物理与计算基础提供了新见解,并为未来意识研究指明了可能方向。
关键词:大脑,意识,熵,知觉,不确定性
1 | 引言 感觉知觉始于一个外部刺激(S),该刺激引发神经反应(R)。刺激本身无法被大脑直接获取,因此成为一个潜在或隐藏变量。因此,大脑所接触到的并非刺激本身,而是其表征(即神经反应)。由于两个原因,R 并非 S 的完美表征:首先,神经反应具有噪声;其次,神经反应所包含的关于刺激的信息不足。神经反应对刺激的不完美表征意味着,对大脑而言,刺激是由反应所欠定的(under-determined)——即无法从反应中无歧义地解码出刺激。结果,大脑对刺激始终存有不确定性。[1]
鉴于感觉数据的不完备性,大脑会进行因果推断,以补充不足的信息,从而降低其对外部刺激的不确定性。[2] 因果推断处于感觉处理核心的观点在当代神经科学中被广泛接受,并构成诸多主流知觉理论的核心组成部分,包括自由能原理[3]和贝叶斯因果推断理论。[4]
本研究提出一种基于不确定性与信息概念的感觉处理与意识知觉模型,此处所用“信息”与“不确定性”均依照香农(Shannon)所建立的信息论中的定义。[5] 在这一点上,本文所提出的意识模型与最具影响力的意识理论之一——整合信息理论(Integrated Information Theory, IIT)形成对比。IIT 所定义的整合信息,是指一个系统整体所产生的、不可还原的、超越其各部分之和的信息,并将此视为意识本身。[6] 在该理论中,信息及其伴随的不确定性之使用方式,与经典信息论中的用法存在显著差异。在 IIT 框架内,信息具有因果性和内在性,其评估源自系统自身的内在视角;相比之下,香农信息则具有观察性和外在性,是从外部观察者的立场进行评估的。[7] 相较于偏离这些标准定义的模型,一个基于标准且被广泛接受的信息与不确定性定义的意识模型,或许更具优势。
2 | 不确定性与熵
2.1 | 不确定性的背景
不确定性是神经计算固有的一部分,无论涉及感觉处理、运动控制还是认知推理。[8] 在感觉处理中,不确定性主要源于两个来源:内部噪声和信息不足。
内部噪声(或称神经噪声)是指神经对刺激的反应中无法由刺激本身解释的部分。内部噪声部分源于随机效应(例如,神经递质释放的随机性或离子通道的随机开闭),部分源于不可控但非随机的效应。[9] 内部噪声通过“神经反应变异性”导致不确定性,即同一刺激在不同试验中可引发不同的神经反应。换句话说,当同一刺激被重复呈现给观察者时,每次都会产生不同的神经活动模式,这些模式针对该刺激的特定特征进行调谐。[10]
当呈现相同的感官变量(如视觉刺激的运动方向)或运动指令时,皮层神经元的放电次数在不同试验间差异很大,通常符合类泊松统计。必须认识到,变异性与不确定性密不可分:若不存在神经变异性——即每当看到同一物体时神经元都以完全相同的方式放电——那么我们对该物体的感知将始终确定无疑。[11] 尽管 Ma 等人[11]正确地将反应变异性识别为不确定性的来源,但应强调的是,反应变异性并非感觉处理中不确定性的唯一来源。另一个来源在于神经对刺激的反应缺乏关于该刺激的充分信息,部分原因是感觉受体无法从外部刺激中提取所有相关信息,部分则源于皮层神经元在处理来自感觉受体的信息时存在固有局限。
Pouget 等人[12]强调了源自内部噪声的不确定性。然而,无论是否存在内部噪声,不确定性本质上都内嵌于大多数相关计算的结构之中。在计算机视觉领域,不确定性长期以来以“病态问题”(ill-posed problems)的形式被研究。关于图像的大多数问题都是病态的,因为图像本身缺乏足以得出无歧义答案的信息。运动处理中的“孔径问题”(aperture problem)便是一个著名例子:当一根棒在孔径后移动时,存在无限多种二维运动与所观察到的图像一致,因此仅凭图像无法确定物体的真实运动。尽管这个例子看似人为(毕竟大多数图像并非孔径后的棒),但它对神经系统提出了真实挑战:所有视觉皮层神经元都是通过其感受野(类似于孔径)观察世界的。此外,类似的问题也出现在计算世界三维结构、定位听觉声源以及其他计算任务中。
事实上,感觉系统必须解决的许多(即便不是全部)问题都是病态的。例如,视觉被认为是病态的,因为视网膜图像可能是视觉场景的任意复杂函数,因此图像中的信息不足以唯一确定该场景。[13] 无论不确定性的来源是噪声还是信息不足,一旦大脑接收到感觉输入,便会启动一个过程,以最小化关于输入原因(即刺激)的不确定性,这构成了感觉知觉的本质。
在涉及感觉处理的心理物理学实验中,不确定性通常使用信息论中的互信息(mutual information)度量进行量化。互信息随神经回路中不确定性(熵)的降低而增加,前提是给定特定的感觉刺激特征。[14] 在视觉或听觉知觉任务的电生理记录中,互信息衡量的是:观察一段脉冲序列(或多个神经元的多段脉冲序列)能在多大程度上减少关于外部刺激的不确定性。[15]
2.2 | 不确定性最小化
在信息论中,不确定性意味着信息的缺失,而最小化不确定性即对应于获取信息(或知识)。在感觉知觉的语境下,不确定性指的是大脑关于外部刺激的信息缺失。在此语境中,不确定性是大脑对刺激“信念”(belief)的一个属性。[1] 此处,“信念”是一个技术术语,指大脑关于刺激的知识。数学上,信念可被视为一个“概率分布”,[16] 其宽度反映了不确定性的程度:概率分布越分散,不确定性就越大。[1]
多项证据支持大脑实际上以概率分布的形式表征不确定性。例如,功能性磁共振成像(fMRI)研究表明,在人类视觉皮层中,感觉不确定性由神经元群体整体活动所编码的概率分布来表征。[17] 另一个例子是,Walker 等人[18]通过对执行任务的猴子视觉皮层进行多电极记录,证明了编码刺激的同一神经元群体也同时编码关于该刺激的不确定性,后者体现为概率分布的宽度。
为检验感觉不确定性是否确实以皮层神经元活动中概率分布的宽度形式进行编码,van Bergen 和 Jehee[19] 在人类视觉皮层开展了一项 fMRI 研究,实验中参与者经历了著名的“序列依赖”(serial dependence)知觉错觉。他们的研究结果直接表明,视觉区 V1、V2 和 V3 中神经元群体活动所产生的概率分布宽度,确实可作为这些区域中感觉不确定性程度的度量。
因此,在感觉处理的每个阶段,同一个概率分布既表征了知识(即信念),也表征了大脑关于刺激的知识缺失(即不确定性)。为了最小化不确定性(从而获取知识),大脑会操作并整合大脑皮层不同神经元群体所编码的概率分布。通过操作和整合概率分布来最小化不确定性,实际上是一种概率推理。若该过程遵循贝叶斯规则,则称为贝叶斯推理。心理物理学实验已证实,大脑在知觉和行动中均执行贝叶斯推理以应对不确定性。[20] 这一观点引出了感觉知觉的贝叶斯方法:大脑作为隐藏刺激(S)的贝叶斯观察者,该刺激产生了神经反应(R)。[21]
2.3 | 通过贝叶斯推理实现不确定性最小化
处理涉及不确定性任务的一种高效方法是通过概率分布表征知识,并通过概率推理获取新知识。[8] 已有研究表明,大脑不仅表征概率分布,还能依据贝叶斯规则对其进行操作和组合。[12]
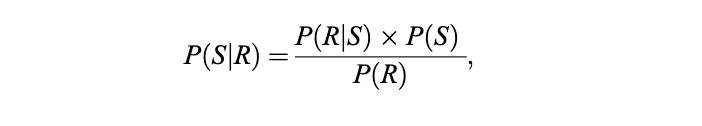
其中 P表示概率,S表示外部刺激,R表示对该刺激的神经反应(即刺激的内部表征)。符号 ∣(竖线)表示条件概率,意为“在……条件下”。
P(S∣R)被称为后验概率分布(或后验信念)。它包含了大脑在考虑神经反应之后关于刺激的全部知识。后验信念不仅反映了刺激的最可能取值,也通过分布的宽度反映了大脑对该刺激的不确定性。[22]
P(R∣S)是反应概率分布,反映了所有可能产生所观察到的反应的不同刺激的似然性。[23] 在层级预测编码(hierarchical predictive coding)框架中,P(R∣S)对应于依赖于下层预测误差的自下而上效应。[24]
P(S)被称为先验信念。更准确地说,它是关于刺激的先验概率分布,包含了大脑在考虑神经反应之前关于该刺激的任何信息。先验信念源于环境的统计规律性。[25] 在层级预测编码框架中,P(S)对应于自上而下的预测。
P(R)是关于反应的先验概率分布,被称为“边缘似然”(marginal likelihood)或“模型证据”(model evidence)。P(R)仅是一个归一化因子,用于确保 P(S∣R)是关于 S的一个合法的概率分布。
在自由能原理(free-energy principle)的框架下,模型证据的负对数被称为“惊奇”(surprise);因此,最大化模型证据等价于最小化惊奇。[26] 从随机热力学(stochastic thermodynamics)的角度看,这种惊奇的时间平均被称为熵或不确定性。[27] 一个旨在最大化其证据的贝叶斯大脑,本质上是在试图最小化其熵。这意味着它违背了热力学第二定律,从而为系统在自然趋向无序的背景下仍能实现自组织提供了一个原理性解释。[28] 因此,最大化模型证据不仅对应于最小化变分自由能,也对应于最小化热力学熵。[29]
贝叶斯公式表明,为了执行贝叶斯推理,大脑需要两个关键要素:第一,对先验和似然函数的表征;第二,一种将这些分布相乘的机制。[12] 似然函数与先验的乘积构成一个联合概率分布,记为 P(R,S),称为生成模型(generative model)。[30]
这可以总结为以下陈述:感觉知觉是通过贝叶斯推理实现的不确定性最小化过程。这一陈述在计算层面和算法层面上解释了知觉,但并未说明知觉在实现层面(implementational level)是如何进行的。神经元究竟是如何实现贝叶斯推理和不确定性最小化的?
2.4 | 不确定性最小化的神经实现
总体而言,我们对大脑如何实现不确定性最小化和贝叶斯推理知之甚少。大脑如何表征概率分布,以及如何将先验知识与似然函数结合起来的机制,目前仍很不清楚。尽管使用 fMRI、EEG、脑磁图(MEG)和颅内记录的实验研究已初步揭示了贝叶斯计算可能发生的脑区,但这些计算在神经元群体层面的具体实现机制仍不明确。利用神经网络模型的研究提供了概念验证,表明此类实现原则上是可能的,但大脑神经网络实际采用的机制仍是未来研究的课题。[4]
信息是减少不确定性的要素,而不确定性意味着信息的缺失。为了在实现层面解释大脑最小化不确定性的机制,我们需要回答以下问题:大脑中表征信息的最基本单元是什么? 对此问题最被广泛接受的答案是:神经系统依赖动作电位(action potential)作为信息与信号传递的核心单元。[31] 在大脑中,信息通过动作电位(即“脉冲”或“spikes”)进行编码。[32] 由于动作电位是一种“全或无”(all-or-nothing)的现象,可被视为一种数字信号,在大脑计算中承载信息。[33]
在基于脉冲的计算中,存在多种表征信息的可能性:单个神经元的脉冲发放率(spike rate)、单个神经元的脉冲时序(spike timing)、单个神经元内部的脉冲相关性,以及神经元群体间的脉冲相关性(例如同步放电)。[34] 实验证据表明,所有这些可能性在大脑中确实都存在,且大脑在不同情境下使用不同类型的脉冲编码方案。例如,脉冲发放率(单位时间内的脉冲数量)通常比脉冲时序或脉冲数量对行为决策的影响更大。[35] 在视网膜中,视觉刺激的出现可通过单个神经节细胞动作电位的精确时序以高精度加以表征。[36]
尽管基于脉冲的计算目前是神经科学的主流观点,但不应忽视的是,神经元可能执行远超脉冲所能允许的计算,因为在神经元内部存在一个巨大且很大程度上尚未被揭示的分子层面计算层。[37] 例如,神经元可以利用细胞内钙离子而非动作电位来表征和处理感觉信息。实验证据明确表明,在对声音刺激的反应中,蟋蟀的听觉神经元通过细胞内钙离子浓度的时间动态进行化学计算。[38] 神经元内化学计算的另一个潜在载体是亚细胞振荡器(subcellular oscillators)。即使是一个由少量耦合的细胞内振荡器组成的小型网络,也可能执行复杂的计算。例如,已有研究表明,仅三个相互作用的振荡器就能以 95% 的准确率识别(或预测)一个随机选取点的颜色。[39]
化学计算是生物学的核心,驱动着从蛋白质合成、细菌生命周期到人类大脑复杂功能等生命过程中的非凡现象。[40] 然而,与基于脉冲的计算不同,大脑中的化学计算尚未被充分探索。我们仍不了解单个神经元内部所执行的化学计算的本质与逻辑。[41]
无论大脑是通过电脉冲还是化学基质进行计算,它都必须在计算层面和算法层面最小化其信息论意义上的不确定性,以获取关于环境的信息。信息论中的“不确定性”在形式上等价于热力学中的“熵”。[42,43] 尽管信息论不确定性与热力学熵衡量的是两种不同性质的量,但它们之间存在如此根本的联系,以至于热力学定律被视为信息论中的定理。[44] 因此,可以合理地认为:在大脑中,计算和算法层面的不确定性最小化,对应于实现层面的热力学熵最小化。换句话说,感觉知觉不仅涉及大脑信息论不确定性的最小化,也涉及其热力学熵的最小化——即承载大脑计算中信息的物理基质的熵的降低。
3 | 熵与相干性
在热力学和统计力学中,熵是无序度的度量,[45,46] 系统的熵越低,其有序性就越高。这一原理适用于任何物理系统,无论该系统的组成要素是动作电位、钙离子还是亚细胞振荡器。原则上,所有这些要素都可以编码信息,并在大脑计算中表征概率分布。正如 Timme 和 Lapish 所指出的,[47] 当一个系统的概率分布偏向某些特定状态时,其熵较低,反映出不确定性减小;相反,当概率在多个状态之间均匀分布时,熵较高,表明不确定性更大。当一个变量完全被限制在单一状态(概率为 1)时,其熵降至 0,表示不存在任何不确定性。
零熵是经典物理学中一个系统所能达到的最小热力学熵。在感觉信息处理的语境下,零热力学熵对应于大脑对外部刺激的不确定性为 0 的状态,即大脑对该刺激完全确定。这是熵(以及不确定性)最小化的终极极限,也等价于大脑在感觉知觉中所能达到的确定性的最高极限。然而,这一极限仅在我们假设大脑仅使用经典信息进行计算的前提下成立,因为量子信息允许达到超越传统确定性的确定性水平。[48]
这种非凡效应源于包含量子信息的纠缠态。纠缠是一种量子资源,它使得系统的熵可以变为负值(即小于零),尽管这在经典物理中是被禁止的。[49] 当一个系统的熵变为负值时,该系统将变得比经典情形下可能达到的有序程度更加有序。这种高度有序的状态被称为量子相干态(或纯态)。量子相干性与量子纠缠密切相关,二者均源于叠加原理——即量子系统能够同时处于多个状态的能力。[50,51]
4 | 相干性与熵
实验证据表明,量子纠缠与相干性可能参与了多种不同的生物现象,包括植物和细菌中的光合作用。[52] 在光合系统 Fenna–Matthews–Olson 复合物中发现的长寿命量子相干性表明,量子相干性可能在生物分子内的能量传递中发挥功能性作用。[50] 除了生物系统中的能量传递外,相干性与纠缠也可能参与鸟类磁罗盘的功能及其导航机制。[53] 意识是另一个可能涉及纠缠与相干性的重要生物现象。[54] 此处需强调的是:虽然量子相干性在光合作用中的作用得到了强有力的实验证据支持,其在鸟类磁罗盘中的参与仅有较弱的实证依据,而其与意识涌现的关联则仍属高度推测性,缺乏实证基础。
近期提出的一种意识理论——量子力学、生物学、信息论与热力学理论(QBIT 理论)——认为,要使感觉处理进展到意识知觉的阶段,量子纠缠与相干性是不可或缺的。[55–57] 根据 QBIT 理论,当大脑关于某一刺激的信息论不确定性降低至某一临界阈值以下时,大脑便对该刺激产生意识。[58–60] 该临界阈值被设定为 0。因此,要使大脑对该刺激产生意识,信息论不确定性(从而热力学熵)必须降至 0 以下(即变为负值)。
5 | 结论
感觉知觉是一个计算过程,大脑通过贝叶斯推理试图最小化其对外部刺激的不确定性。在此过程中,根据不确定性被最小化的程度,刺激会被大脑有意识地或无意识地感知。只有当信息论不确定性降低至某一临界阈值以下时,才会发生有意识的知觉。这是一个极端阈值,超出了经典资源(如能量)所能达到的极限。要产生意识,大脑需要量子资源,包括纠缠。
意识的涌现不仅与计算层面信息论不确定性的降低相关,也与承载(或编码)信息的物理基质在实现层面热力学熵的降低相关。这一物理基质目前尚不清楚,尽管细胞脉冲和亚细胞振荡器是颇具前景的候选者。熵的降低对应于有序性的增加,而量子相干性是物理基质所能达到的最大可能有序状态。[57] 本文提出:当大脑对某一刺激产生意识时,其不确定性以及计算基质的熵会变为负值。如果计算基质是脉冲,则意识的涌现与全脑神经元的相干放电相关;如果基质是化学振荡器,则意识的涌现与全脑亚细胞实体的相干振荡相关。无论如何,全脑的相干活动是意识的标志性特征(见图 1)。
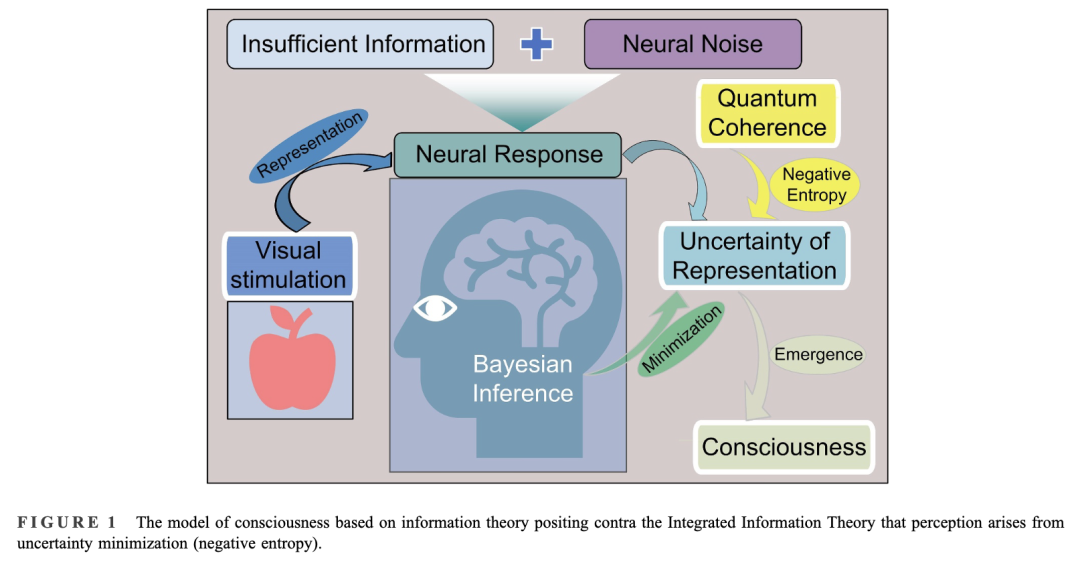
QBIT 理论面临的一个挑战是目前尚无任何直接的实验证据支持;它完全是推测性的。此前一篇论文[56]提出一个相关概念,旨在应对这一挑战,即熵最小化与热量移除之间的关系。将熵从系统中导出,等同于从系统中移除热量,从而降低系统温度,使系统变得更冷。因此,为实现意识涌现所必需的极端不确定性(熵)最小化所需的神经计算,不仅不会产生热量,反而会移除热量,从而冷却其底层的神经介质。这表明,意识可能充当大脑的一种冷却机制——这一假说可通过专门设计用于测量产生意识体验的神经回路温度的仪器加以验证或证伪。
原文链接:https://www.biorxiv.org/content/10.1101/2025.06.25.661321v1.abstract
本文参与 腾讯云自媒体同步曝光计划,分享自微信公众号。
原始发表:2025-10-18,如有侵权请联系 cloudcommunity@tencent.com 删除
评论
登录后参与评论
推荐阅读
腾讯云开发者

Copyright © 2013 - 2026 Tencent Cloud. All Rights Reserved. 腾讯云 版权所有
深圳市腾讯计算机系统有限公司 ICP备案/许可证号:粤B2-20090059 ![]() 粤公网安备44030502008569号
粤公网安备44030502008569号
腾讯云计算(北京)有限责任公司 京ICP证150476号 | 京ICP备11018762号