【免疫学实验】单克隆抗体技术

【免疫学实验】单克隆抗体技术

三兔测序学社
发布于 2025-12-29 12:55:40
发布于 2025-12-29 12:55:40
1. 抗体的异质性及其局限
自然或人工免疫产生的抗体,通常是多种不同特异性和亲和力抗体的混合物。
原因: 即使是针对单一抗原决定簇(如半抗原)产生的抗体,其结构也存在差异(异质性)。这可以通过等电聚焦实验验证:识别相同抗原的抗体在电泳后会显示出不同的等电点,证明其分子结构的不均一性。
局限性:
批次差异: 即使使用相同方法免疫基因相同的动物,所得抗血清也各不相同。
产量有限: 无法满足长期、大规模的实验或临床需求。
交叉反应: 即便经过纯化,复杂的抗体混合物仍可能导致非特异性的交叉反应。
2. 单克隆抗体的诞生
为了解决上述问题,需要一种能无限供应、结构均一的特异性抗体。早期生化学家研究多发性骨髓瘤患者的骨髓瘤蛋白,证实了永生化浆细胞能产生单一抗体,但其抗原特异性大多不明。
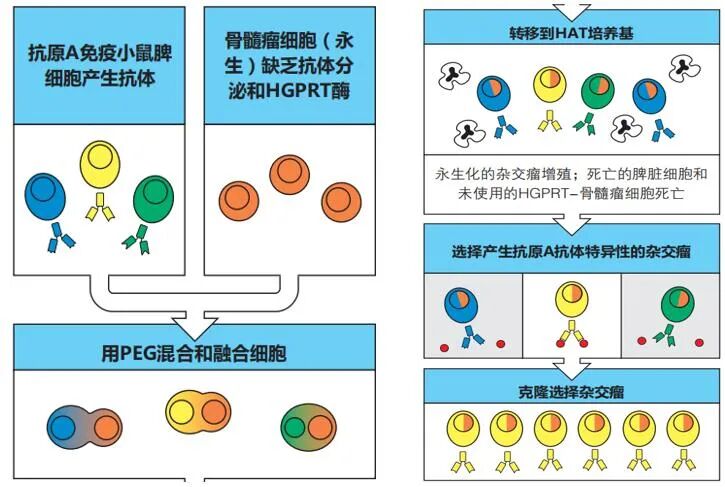
杂交瘤技术的突破:Georges Köhler 和 César Milstein 发明了细胞融合技术:
融合: 将小鼠脾脏细胞(提供特异性抗体产生能力)与骨髓瘤细胞(提供无限增殖能力)融合。
筛选: 利用特定药物筛选出杂交瘤细胞(亲本骨髓瘤细胞被杀死,脾细胞自然死亡)。
克隆: 通过单细胞培养,筛选出能分泌特定抗体的杂交瘤克隆。
由此产生的单克隆抗体,所有分子结构完全相同,具有高度特异性和均一性,彻底打破了传统抗血清的限制。
3. 人源单克隆抗体的现代制备
虽然杂交瘤技术主要应用于小鼠,但目前治疗性的人源单克隆抗体主要通过以下基因工程技术制备:
噬菌体展示技术
重组 DNA 技术(克隆人浆细胞抗体基因)
携带人类抗体基因的转基因小鼠
噬菌体展示技术
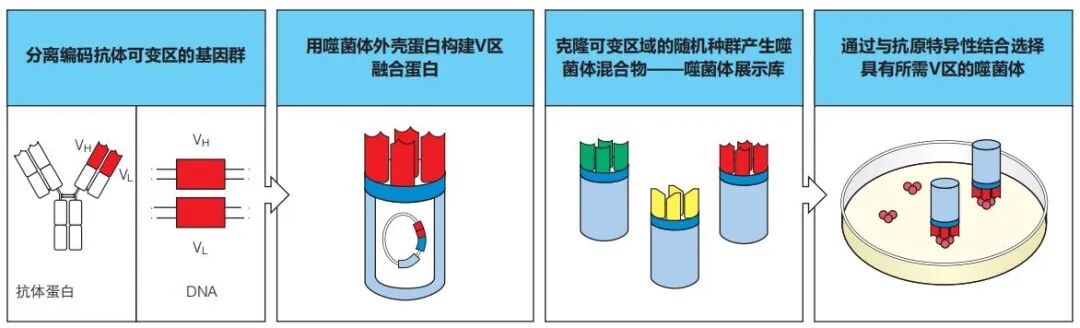
噬菌体展示技术是将编码抗体可变区(V 区)的基因片段与编码噬菌体外壳蛋白的基因融合以产生抗体样分子的方法。将含有这种融合基因的噬菌体感染细菌,所产生的噬菌体颗粒表达抗体样融合蛋白的外壳,而抗体的可变区(抗原结合域)表达在噬菌体的外部。每一个噬菌体都在表面表达不同的抗原结合域,一系列这样的噬菌体总和起来被称为噬菌体展示库。可以用亲和层析法)分离在噬菌体展示库中与特定抗原结合的噬菌体,所分离的噬菌体颗粒可用于感染新鲜细菌。以这种方式分离的每个噬菌体都可以产生类似于单克隆抗体的单克隆抗原结合颗粒(图 2)。
每个噬菌体都编码独有的抗原结合位点的基因,并可从噬菌体 DNA 中将其恢复出来,通过将其连接到编码抗体恒定区的免疫球蛋白基因来构建完整抗体分子的基因。当这些重组抗体基因被导入合适的宿主细胞系,例如导入非抗体产生的杂交瘤骨髓瘤细胞时,被导入的细胞就可以分泌针对特定抗原的单克隆抗体。
从接种过疫苗的个体中产生人单克隆抗体:由于接种过疫苗的个体所产生的抗体已经重排重链和轻链基因序列,所以在某些情况下,可以从这些个体中分离浆细胞来制备人单克隆抗体。人血浆细胞可以来自约 1 周前受接种个体的外周血,根据细胞表面分子(如 CD27 和 CD38)将单个浆细胞分离到微量板中,用PCR 从每个细胞中克隆抗体重链和轻链可变区序列。然后将这些序列插入克隆载体以重建全长的抗体重链和轻链基因,再将成对的重链和轻链载体导入永生化的人类细胞系,最终筛选出分泌与免疫抗原相结合的抗体的细胞系,作为特定人源抗体的永久来源。
本文参与 腾讯云自媒体同步曝光计划,分享自微信公众号。
原始发表:2025-12-16,如有侵权请联系 cloudcommunity@tencent.com 删除
评论
登录后参与评论
推荐阅读
腾讯云开发者

Copyright © 2013 - 2026 Tencent Cloud. All Rights Reserved. 腾讯云 版权所有
深圳市腾讯计算机系统有限公司 ICP备案/许可证号:粤B2-20090059 ![]() 粤公网安备44030502008569号
粤公网安备44030502008569号
腾讯云计算(北京)有限责任公司 京ICP证150476号 | 京ICP备11018762号